.png)

Main
The origin of tetrapods, understood as an evolutionary and ecological phenomenon, was not a single event but a process that began with the acquisition of incipient terrestrial locomotory competence in the tetrapod stem group and ended with the emergence of the major crown-group clades, amphibians and amniotes. Of particular importance for the future development of the global ecosystem was the origin of amniotes, the only tetrapod clade to achieve complete reproductive independence of water, and by far the most impactful in terms of both diversity and disparity.
An overall understanding of this phase of vertebrate evolution requires data on phenotypic change, the timing of evolutionary and cladogenetic events, and patterns of diversity, disparity and biogeography. Three principal data sources are available: body fossils, ichnofossils (footprints and other traces) and time-calibrated molecular phylogenetic divergence dates. Body fossils and ichnofossils are typically preserved in different sedimentation regimes, and can thus capture animals with different environmental preferences, but both require taphonomic settings with net sediment deposition rather than erosion, and will thus be biased towards lowland environments, although some upland depositional settings are also known10. Molecular divergence dates are unaffected by depositional environments, but are themselves partly dependent on fossil calibration of the phylogeny. Furthermore, they can date only phylogenetic nodes uniting living lineages, such as the tetrapod crown-group node (uniting the lissamphibian and amniote lineages) and the amniote crown-group node (uniting the mammal and reptile–bird lineages). Fossils, by contrast, can illuminate the details of morphological evolution within stem groups.
Molecular divergence dates for the amniote crown-group node from 30 recent studies (Supplementary Information Part 1), curated at the TimeTree website (https://www.timetree.org), form a tight cluster with a median age of 319 million years, which corresponds to early Bashkirian (mid-Carboniferous); the spread of the cluster is 308.5 to 334.7 million years, thus spanning from Moscovian (late Carboniferous) to Viséan (early Carboniferous). The corresponding date cluster of 32 dates for the tetrapod crown-group node has a much wider spread, ranging from 333.3 to 395.0 million years (that is, from the Viséan to the Eifelian (Middle Devonian)); the median age in this case is 352 million years, or Tournaisian (earliest Carboniferous). The preponderance of molecular evidence thus suggests an origin of the tetrapod crown group during the earliest Carboniferous, with crown amniotes appearing some 30–35 million years later. This places these events in the aftermath of the end-Devonian mass extinction, during and after the 20-million-year interval of poor fossil record known as Romer’s gap11. The published fossil record is compatible with this time frame, showing the earliest crown-group amniote body fossils and trackways (Hylonomus and Notalacerta) in the Bashkirian1,2,3,4, the earliest crown-group tetrapod body fossils (for example, Balanerpeton) in the late Viséan5,6, and the earliest crown-group tetrapod trackways (for example, Batrachichnus and Palaeosauropus) in the mid-Tournaisian7 (Fig. 1a). However, this compatibility partly reflects the calibration of the molecular trees by known fossils, and is thus not a fully independent verification.
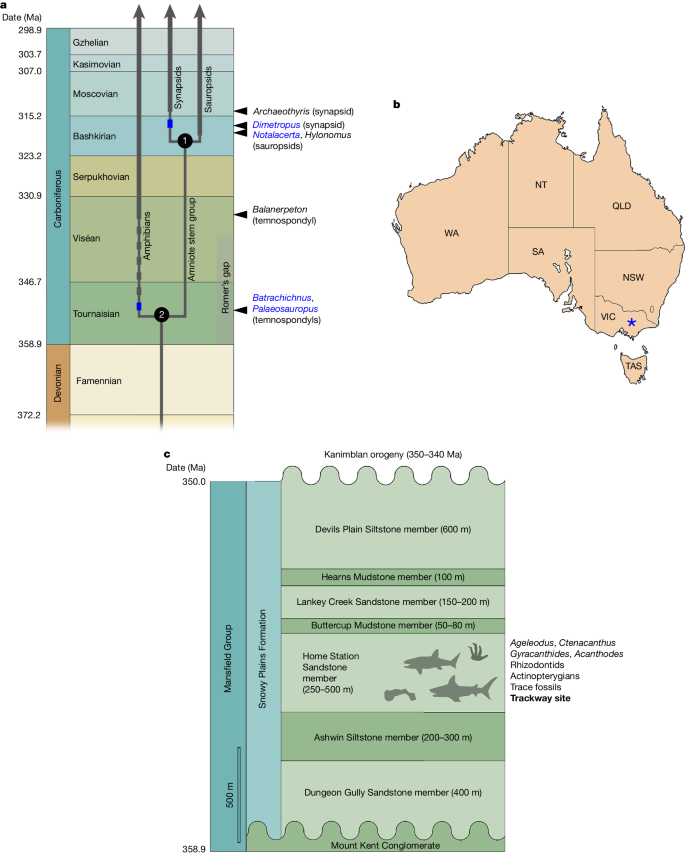
a, Stratigraphic timescale representation of the known early fossil record of crown-group tetrapods. Thin grey lines indicate phylogenetic branches; thick grey lines indicate the body-fossil record from the earliest occurrence; arrowhead and name in black on the right margin indicate the name of the earliest body fossil; blue rectangles indicate the earliest ichnofossil record when this is earlier than the body-fossil record; the dashed line of grey rectangles indicates the range extension between the earliest body fossil and the earliest ichnofossil; name in blue on the right margin indicates the name of the earliest ichnorecord. The amniote crown-group node (1) and tetrapod crown-group node (2) are given minimum ages compatible with the fossil record. All dates are from https://stratigraphy.org/chart. Ma, million years ago. b, Map of Australia showing the locality (blue asterisk). NSW, New South Wales; NT, Northern Territory; QLD, Queensland; SA, South Australia; TAS, Tasmania; VIC, Victoria; WA, Western Australia. c, Stratigraphy of the Mansfield Group.
We present here new trackway evidence from Taungurung Country, Victoria, Australia (Figs. 1b,c and 2), indicating that these dates are substantially too late. Crown amniotes were already present in northeast Gondwana by the early Tournaisian. This in turn implies that the crown tetrapod node must lie deep in the Devonian. New trackway data from Silesia in Poland show that the earliest records of crown amniotes in the equatorial regions of Euramerica are also earlier than previously thought, Serpukhovian rather than Bashkirian.

a, Photo of the slab, NMV P258240, as preserved. b, Same as in a, with footprints and trackways highlighted. Manus (front foot) prints are shown in yellow; pes (hind foot) prints are shown in blue. Am1–4, manus prints from trackway A; Ap1–4, pes prints from trackway A; Bm1–5, manus prints from trackway B; Bp1–4, pes prints from trackway B; Ip, isolated right pes print. c,d, Isolated right pes print Ip as a false-colour inverted scan image (c) and photo (d). e,f, Right manus print Am1 as a false-colour scan image (e) and photo (f). g, Photo of pes print Bp4 (above) and manus print Bm3 (below). In c–g, white arrows denote claw impressions or scratches, Roman numerals denote digit numbers. Scale bars, 50 mm (a) and 10 mm (c–g).
The Australian tracks
The Australian tracks are preserved on the upper surface of a loose but essentially in situ fine-grained silty sandstone block from the bank of the Broken River at Barjarg, Taungurung Country, Victoria (Museums Victoria specimen NMV P258240). In the Taungurung language, this section of the Broken River was referred to as Berrepit, meaning to flee or run away12. The block derives from the Home Station Sandstone member of the Snowy Plains Formation in the upper part of the Mansfield Group (Fig. 1b,c). Although aquatic invertebrate and fish trail trace fossils have previously been described from other locations within the Home Station Sandstone13, this is the first record of terrestrial vertebrate tracks. This new specimen was discovered by the two non-professional members of the author group (C.A.E. and J.E.), who brought it to the attention of the professional palaeontologists, in a demonstration of the value of citizen science. The locality lies within the Lachlan Fold Belt, a tectonically complex region with multiple sedimentary basins and several orogenic episodes during the Palaeozoic (Supplementary Information Part 2). The sedimentary deposits in the region are predominantly Devonian and include several important vertebrate localities8. However, the upper Mansfield Group9 contains a characteristic early Carboniferous vertebrate assemblage without Devonian index taxa such as placoderms, tristichopterids and porolepiforms14,15,16,17. An associated assemblage of vertebrate microremains has yielded a tooth of the chondrichthyan genus Ageleodus, which closely resembles teeth of this genus from the Famennian (latest Devonian) of the Catskill Formation in Pennsylvania, rather than examples from later Carboniferous localities14. Deposition in this region was terminated by the Kanimblan orogeny, which began in the latest Devonian and during which active folding seems to have come to an end by the late Tournaisian18,19 (Supplementary Information Part 2). The Snowy Plains Formation must thus belong to the early part of the Tournaisian; it probably falls within the age span 358.9 to 354 million years old.
The track surface, which is dense, fine-grained and very well preserved, carries three generations of subaerial original surface tracks, preserved in concave epirelief, which all seem to have been made by the same trackmaker taxon (Fig. 2a,b). The oldest is an isolated pes print (Ip). A brief rain shower after this footprint had been made left it, as well as the general surface, pockmarked with raindrop prints. Shortly after the rainfall, while the ground was still moist, a trackway (A) was made by an animal that left well-defined foot impressions. Sometime later, when the ground had begun to dry and harden, another trackway was made (B) that consists largely of well-preserved claw scratches with faint accompanying footprints. Neither trackway is associated with a body or tail drag. The spacing of manus and pes prints in trackway A implies a hip–shoulder distance of approximately 17 cm if the animal was trotting, slightly more if it was performing a sequential walking gait. The total body length is impossible to determine because neck plus head length and tail length are unknown, but applying the proportions of a modern water monitor (Varanus salvator), which has a broadly similar foot morphology, gives a suggested length in the region of 80 cm.
The tracks present a consistent foot morphology, well documented by the combined evidence of the different footprints (Supplementary Information Part 3). The manus is smaller than the pes (Fig. 2g). Both are pentadactyl, with five relatively long, slender digits splayed out into a fan shape, although digit V does not always leave a distinct impression. Digits I and V are the shortest, III and IV the longest. The digit impressions bulge distally into slightly swollen tips, but there are no distinct phalangeal pad impressions (Fig. 2e,f). The impression of the skin surface appears smooth without distinct scales, although this may be a preservation effect. Digits I–IV are associated with impressions of fairly large sharp claws, similar in relative size to those of a monitor lizard (Varanus), whereas digit V carries a very short claw. The claw prints are sometimes deflected medially at approximately 90° to the long axes of the digits, creating a characteristic inverted J shape (Fig. 2c,d). The claws are discrete structures, clearly distinct from the more proximal parts of the digits, as shown by the co-occurrence of softly rounded digit tips with sharp claw scratches in the same footprint (Fig. 2e,f).
This foot morphology carries a clear phylogenetic signal. Claws are a derived character of crown amniotes and are almost invariably present in this clade. Importantly, they are not present in known stem amniotes; seymouriamorphs, diadectids and limnoscelids all lack claws, as evidenced by their footprints7,20,21,22,23,24,25. As claws must have been present at the amniote crown-group node, it is probable that they originated at the very top of the stem group, but no unambiguous clawed stem amniotes have been discovered. Claws were apparently present in microsaurs10, which have an uncertain phylogenetic position but may be crown amniotes6. Outside the crown amniotes, claws or keratinized toe tips occur sporadically in modern anurans (Xenopus and Hymenochirus)26 and salamanders (for example, Onychodactylus), and may have been present in some temnospondyls judging by the shape of the terminal phalanges27. However, ichnotaxa such as Batrachichnus and Limnopus, which are attributed to temnospondyls, lack claw impressions7, and in any case, the presence of five digit-impressions on the manus prints of the Snowy Plains Formation slab rules out a temnospondyl identity for our trackmaker.
Within the amniote crown group, the deepest phylogenetic split is that between Synapsida (stem and crown mammals) and Sauropsida (stem and crown reptiles, including birds). The earliest known synapsid ichnogenus is Dimetropus, thought to represent ‘pelycosaur’-grade stem mammals, which is first recorded in the Bochum Formation (late Bashkirian) of Germany24. The earliest described sauropsid ichnotaxon is Notalacerta, which has been described from mid-Bashkirian localities. The broadly similar Varanopus and Dromopus are slightly younger2,25. These tracks have been ascribed to stem reptiles including captorhinids, protorothyrids and basal diapsids2,7,25,
Dimetropus prints are clearly different from those on the Snowy Plains Formation slab, notably in having longer soles with distinct ‘heels’, less splayed toes and longer, straighter claws (Fig. 3h). By contrast, the Snowy Plains Formation tracks closely resemble Notalacerta, Dromopus and in particular Varanopus (Fig. 3a–c,e–g). All of these ichnotaxa have a short sole, which often leaves no impression, and digit impressions splayed into a fan shape. They are ectaxonic, meaning that the lateral digits are more strongly developed than the medial ones, and they all show claw impressions, which are not just distinctively sharp-pointed but are also commonly deflected medially (towards the body of the animal) from the long axis of the digit print to create J- or L-shaped digit impressions1,2. This exact effect is also seen in the Snowy Plains Formation tracks (Fig. 2); we conclude that the toes of all these trackmakers bore similar claws that were clearly offset from the more proximal part of the digit.

a–c, Three footprints of Notalacerta from the middle Serpukhovian to early Bashkirian Wałbrzych Formation of Silesia, Poland; each is shown as an optical scan (top) and photo (bottom). Holy Cross Branch of the Polish Geological Institute – National Research Institute in Kielce, Muz. PGI-OS 220/182 (a), 184 (b) and 185 (c). d, Isolated left pes print Ip from the Snowy Plains Formation slab NMV P258240 (Fig. 2c,d), reproduced here to facilitate comparison with other amniote footprints. e–g, Presumed sauropsid prints, manus (top) and pes (bottom), of Notalacerta (e), Varanopus (f) and Dromopus (g), all from ref. 2. h, Dimetropus manus or pes imprint, natural cast, IGWU-1, Geological Museum of the Institute of Geological Sciences, University of Wrocław, Wrocław. Labelling as in Fig. 2. Scale bars, 10 mm. Photos in e–g reproduced from ref. 2, Frontiers Media, under a Creative Commons licence CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/).
Notwithstanding these similarities, the Snowy Plains Formation tracks also bear some resemblance to another ichnotaxon, Hylopus hardingi, which is not attributed to crown amniotes. The presumed Hylopus trackmaker is often referred to as a reptiliomorph2, an imprecise term that is best interpreted as equivalent to stem amniote, although phylogenetic instability near the tetrapod crown-group node means that several groups of early tetrapods that have been considered as putative Hylopus trackmakers are retrieved either as stem amniotes or as stem tetrapods at present depending on the particular analysis2,6. Comparison between the Snowy Plains Formation tracks and Hylopus is made more complicated by the fact that the tracks attributed to Hylopus vary in morphology and include examples that appear to have claw impressions2, suggesting that some tracks have been misattributed to this ichnotaxon and could themselves represent early unrecognized sauropsids. However, typical and well-preserved H. hardingi footprints have distinctive, rounded (almost ball-shaped) toe-tip impressions without any trace of claw marks, and a very short digit V on the manus2. The Snowy Plains Formation footprints with their unambiguous discrete claw marks and long digit V on the manus cannot be attributed to H. hardingi.
This character distribution allows for two possible phylogenetic interpretations. There is general agreement that Notalacerta, Dromopus and Varanopus are sauropsid ichnotaxa, but whereas the foot morphology at the amniote crown-group node must have been pentadactyl and claw-bearing, the exact shape of the foot at this node is not known. The overall shape similarity between these ichnotaxa and H. hardingi may reflect a foot morphology conserved across the amniote crown-group node. The occurrence of some Hylopus-like traits in the Snowy Plains Formation tracks suggests either that the trackmaker was a very primitive sauropsid, phylogenetically basal to the Varanopus, Notalacerta and Dromopus trackmakers, or that it occupied a position close to the amniote crown-group node. In the first case, the crown-group node must predate this trackway slab; in the second, it could be contemporary with it. A substantially younger date for the crown-group node is ruled out, unless the crown amniote characters of the Snowy Plains Formation tracks are dismissed (with no evidential basis) as convergences.
The Silesian tracks
The earliest crown amniote fossils acknowledged in the current literature are trackways of Notalacerta and associated body fossils of the stem reptile Hylonomus (which may have been the trackmaker) in the middle Bashkirian of Joggins, Nova Scotia, Canada1,2. However, during this study, tracks similar to those of Notalacerta have also been discovered in the Wałbrzych Formation of the Intra-Sudetic Basin of Silesia in Poland, which has been dated as mid-Serpukhovian to early Bashkirian (Namurian A) on the basis of palynostratigaphy28 (Fig. 3a–c). This pushes back the amniote record of Euramerica by approximately eight million years. The Carboniferous–Permian sites of the Intra-Sudetic Basin (Czech Republic and Poland) are historically important for the study of tetrapod tracks and have been studied for more than 150 years29. Rich material collected from the Carboniferous part of this succession provides a new guide for resolving the first appearance, diversification and environments of early amniotes in this part of Euramerica (Supplementary Information Part 4).
Discussion
The implications of the Snowy Plains Formation trackways are profound and wide-ranging (Fig. 4). In terms of tetrapod evolution, the Devonian Period has, until now, been seen as the exclusive domain of the stem group. All recent palaeontological analyses place the lungfish–tetrapod node close to the Silurian/Devonian boundary30,31,32. The earliest known fish member of the tetrapod stem group, Tungsenia, is Pragian31. Limbed stem tetrapods appear in the ichnorecord in the Eifelian, and in the body-fossil record in the Frasnian33,34,35,36,37. The known tetrapod body-fossil record of the Frasnian and Famennian consists exclusively of stem-group forms. This fits with the idea of the crown-group radiation as a post-Devonian phenomenon following (and possibly impelled by) the end-Devonian extinction event10,37. However, the Snowy Plains Formation trackways challenge this interpretation.
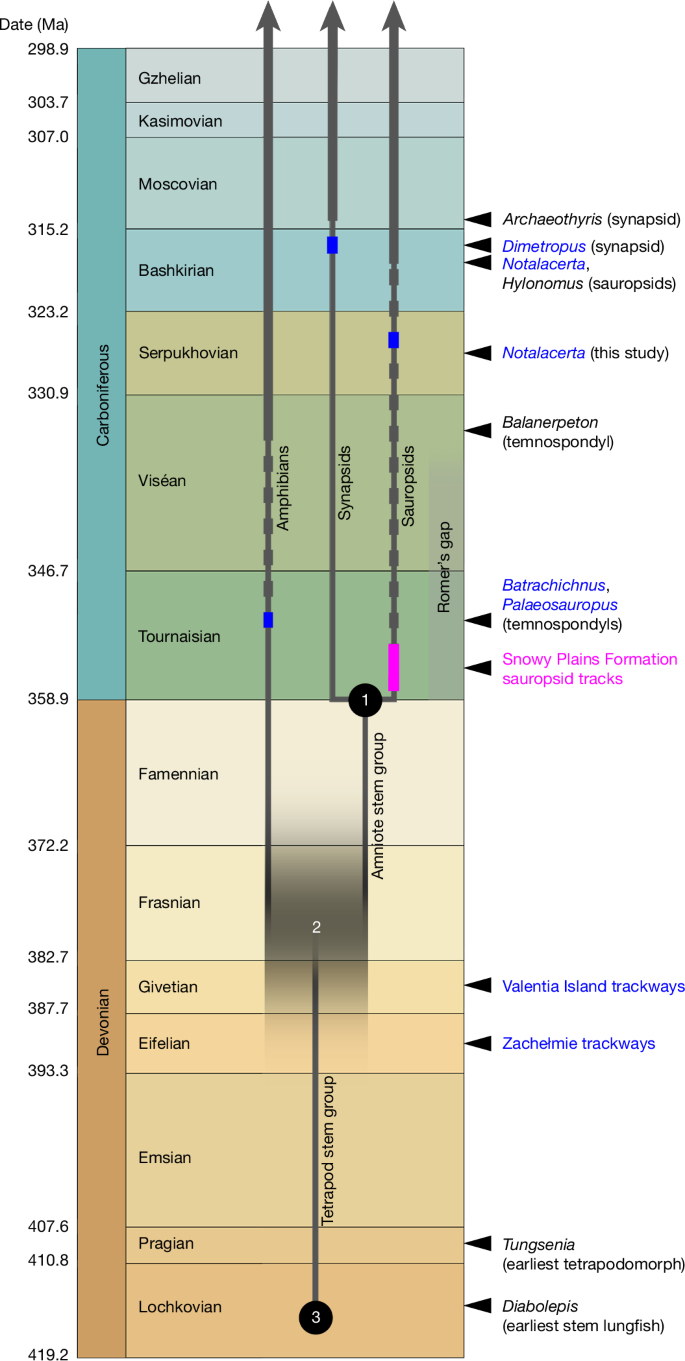
Stratigraphic timescale representation of the Devonian and Carboniferous, showing the impact of the Snowy Plains Formation sauropsid tracks. The track record is shown as a pink rectangle, of double height to indicate possible age range. Other graphic conventions as in Fig. 1. The amniote crown-group node (1) and lungfish–tetrapod node (3) are given minimum ages compatible with the fossil record. The tetrapod crown-group node (2) is positioned in accordance with the branch-length proportions derived from TimeTree (https://www.timetree.org) as explained in the text; vertical blurring of the horizontal branch segment indicates that this date is uncertain and should be considered only as a general indicator, not a precise estimate.
A substantial time interval between the tetrapod and amniote crown-group nodes is a universal feature of recent molecular phylogenies in which both nodes are defined (Supplementary Information Part 1); the median age difference between these nodes in the calibrated phylogenies curated by TimeTree is 33 million years. These age differences are inferred from branch lengths that in turn reflect base substitutions recorded in the genomes of extant animals, and are thus not susceptible to the distorting effects of gaps in the fossil record. Even though the exact ages vary between phylogenies, depending both on the fossil calibrations and phylogenetic algorithms used, the substantial age separation between the amniote and tetrapod crown-group nodes is consistent and must be real; these cladogenetic events were separated by tens of millions of years.
As the Snowy Plains Formation is early Tournaisian in age, a sauropsid identification of the tracks implies that the amniote crown-group node can, at a minimum, be only marginally younger than the Devonian/Carboniferous boundary. This in turn means that the tetrapod crown-group node must lie much further back in the Devonian. To arrive at a rough age estimate for the tetrapod crown-group node, one avenue is to look at the lungfish–tetrapod node, which marks the bottom end of the tetrapod stem group. The median TimeTree age for this node is 408 million years. The corresponding ages of the amniote and tetrapod crown-group nodes are 319 and 352 million years; if the time distance from the amniote crown-group node to the lungfish–tetrapod node is given the unit value 1, the corresponding distance from the amniote crown-group node to the tetrapod crown-group node is 0.371, and that from the tetrapod crown-group node to the lungfish–tetrapod node is 0.629.
In fact, the earliest unambiguous stem lungfish, Diabolepis, is approximately 415 million years old (Xitun Formation, Lochkovian, China)38, so the inferred date for the lungfish–tetrapod node is slightly too young. If, as a thought experiment, the amniote crown-group node and the lungfish–tetrapod node are fixed, respectively, to the Devonian/Carboniferous boundary (358.9 million years) and the mid-Lochkovian (415 million years), and the aforementioned relative branch lengths are applied, they place the tetrapod crown-group node at a median age of 379.7 million years (early Frasnian). This should be understood only as the approximate mid-point of a wide zone of possibility (Fig. 4). However, a much younger age, at or close to the Devonian/Carboniferous boundary, can be rejected because the internode to the amniote crown-group node becomes implausibly short and incompatible with the substantial branch lengths consistently recovered by molecular phylogenies (Supplementary Information Part 1). Conversely, as neither the amniote crown-group node nor the lungfish–tetrapod node has a constrained maximum age, all three nodes could in fact be considerably older than indicated. The rapidly expanding number of sequenced vertebrate genomes creates potential for more robust future phylogenetic analyses that, with the inclusion of the Snowy Plains Formation tracks as a calibration point, can provide a more precise estimate of the tetrapod crown-group node date.
The earliest fossils of limbed tetrapods, the trackways from Zachełmie in Poland (Eifelian)29 and Valentia Island in Ireland (Givetian)30, are, respectively, about 390 and 385 million years old, and thus compatible with this new inferred timeline (Fig. 4). However, the widely accepted picture of Devonian tetrapods as a low-diversity array of primitive fish-like forms10,37 must be false. The cladogenetic event that gave rise to the tetrapod crown group was preceded by a series of others that gave off the various clades of limbed stem tetrapods, such as baphetids, colosteids and ichthyostegids, and before that the elpistostegalians and various tetrapodomorph fishes6,37. All of these cladogenetic events must now be fitted into, approximately, the first two-thirds of the Devonian period. The origins of stem amniote lineages such as seymouriamorphs and diadectomorphs must lie in the Late Devonian. Remarkably, the inferred age of the tetrapod crown-group node presented here is approximately contemporary with the elpistostegalians Elpistostege and Tiktaalik, often perceived as antecedents and potential ancestors of tetrapods39,40,41,42. This result strongly supports the much earlier origin of limbed tetrapods indicated by the Middle Devonian trackway record, and implies that tetrapods underwent a far faster process of cladogenesis and morphological evolution during the Devonian than has hitherto been recognized.
A series of body-fossil discoveries over the past four decades lend indirect support to this contention, by providing evidence for previously unsuspected diversity and morphological disparity among Devonian tetrapods43,44,45,46,47. Particularly noteworthy is the fact that each new tetrapod locality has yielded one or more new tetrapods, a marked contrast with the wide distribution of associated fishes such as Holoptychius and Bothriolepis, and a sign that our discoveries are sampling a high-diversity global tetrapod fauna with small geographic ranges for individual genera. Nevertheless, the complete absence so far in the Devonian body-fossil record of any crown-group tetrapods, or crownward stem-group clades such as colosteids and baphetids, indicates that this record markedly under-samples the living diversity.
The trackway record casts some additional light on this phenomenon. At present, the oldest records of amphibians, synapsids, sauropsids and limbed stem tetrapods are all ichnorecords7,24,33,34 (Fig. 4). It is well known from later parts of the vertebrate fossil record that trackway assemblages often capture taxa that are not seen in associated body-fossil assemblages48, and this also applies to the Devonian and Carboniferous record. The earliest known high-diversity tetrapod trackway assemblage, from the mid-Tournaisian of Blue Beach, Canada, contains taxa that are not represented among the associated body fossils (for example, temnospondyls)7. The Mansfield Group contains no known tetrapod body fossils14,15,16,17. This is also the case for the two Middle Devonian formations that contain published tetrapod tracks, the Givetian Valentia Slate Formation of Valentia Island, Ireland, and the Eifelian Wojciechowice Formation of Zachełmie, Poland; the former yields only fish49,50, the latter no body fossils at all. The trackway record thus provides direct evidence of the incompleteness of the body-fossil record, and in turn has a key part to play in fleshing out the picture of early tetrapod diversity, even though it is also quite meagre.
The poor fossil record hampers the search for temporal and spatial patterns of distribution. With the discovery of the Snowy Plains Formation tracks, the crown-group amniote record of northeastern Gondwana now predates that of Euramerica by about 30 million years, but we cannot rule out that earlier representatives may eventually be found in Euramerica as well. The mid- to late-Serpukhovian amniote tracks from Silesia are morphologically advanced, with pronounced claws and narrow, elongated digits, and some are quite large (Fig. 3c); this strongly suggests that the evolutionary history of this group is nested deeper in time but not yet recognized in this region (Supplementary Information Part 4). With regard to biogeography and living environments, recent palaeomagnetic reanalysis of the Devonian-Carboniferous pole path of Australia51,52 has revealed that the continent was located much further north during the Famennian to Viséan than had previously been thought. At the time of deposition of the Mansfield Group, the trackway locality lay at a latitude of approximately 17° south, at the southern edge of the tropics. This is quite similar to the equatorial latitude of the Euramerican Notalacerta localities, and does not present a strong case for a distinction between temperate and tropical faunas being a factor in early amniote distribution.
By contrast, the Snowy Plains Formation trackways do cast substantial new light on the effect of the end-Devonian mass extinction event on tetrapod evolution. Until recently53,54,55, the tetrapod fossil record showed a hiatus of approximately 20 million years, known as Romer’s gap, between the end-Famennian and the late Viséan. The pre-gap and post-gap tetrapods appeared substantially different in character, with the post-gap forms showing much higher diversity and disparity, as well as being more advanced and including crown-group tetrapods in their ranks10,37. This gave rise to the idea that the extinction event had served as a reset for tetrapod evolution, allowing the emergence of more modern groups. It has also been linked to terrestrialization after a supposedly aquatic phase of tetrapod evolution in the Devonian10,37. This somewhat simplistic conception of Romer’s gap and its relationship to tetrapod evolutionary history can now be replaced by a more nuanced interpretation. The presence of sauropsid tracks in the early Tournaisian implies that the tetrapod crown-group radiation was well under way in the Late Devonian, and that not only lineages such as temnospondyls, seymouriamorphs and diadectomorphs but also crownward stem tetrapods such as baphetids and colosteids crossed the Devonian/Carboniferous boundary. If this is correct, the mass extinction did not have a role in the emergence of these derived lineages, although it is still possible that the amniote crown group arose in its immediate aftermath. The impact of the extinction on diversity, and especially on the selective removal of archaic tetrapod lineages, is harder to assess but may have been substantial. With the exception of Tulerpeton56 and Brittagnathus47 in the Devonian, and some possible Devonian-grade tetrapods in the Tournaisian53,54,55, all known Devonian tetrapods seem to represent a less crownward segment of the tetrapod stem than any post-Devonian forms6. This suggests a selective extinction with appreciable effects on ecosystem structure.
The Snowy Plains Formation trackways have a disproportionate impact on our understanding of early tetrapod evolution because of their combination of diagnostic amniote characteristics and early, securely constrained date. They demonstrate, once more, the extraordinary importance of happenstance and serendipity in the study of severely under-sampled parts of the fossil record. Against this background, two things stand out: first, that the interpretation of such a fossil record is critically dependent on phylogenetic inferences and cannot be ‘read’ as a literal account of the history of a group; and second, the fundamental, continuing importance of palaeontological fieldwork as a source of new knowledge.
Methods
Specimens were photographed under oblique lighting to emphasize the footprints. Optical scans were undertaken with a RangeVision Spectrum and the resulting STL files were rendered in RangeVision 3D studio 2022.1 for greyscale images or ParaView 5.10.1 for false-colour height maps.
Inclusion and ethics
This paper describes Australian and Polish fossil material, deposited with public museums in those countries (Museums Victoria; Holy Cross Branch of the Polish Geological Institute – National Research Institute in Kielce; Geological Museum of the Institute of Geological Sciences, University of Wrocław). The authors include Australian (J.A.L., J.G., A.M.C. and A.B.C.) and Polish (G.N.) researchers, as well as the two discoverers of the Australian trackway slab (C.A.E. and J.E.). As the Australian specimen comes from Taungurung Country, we have consulted with Taungurung Elder and language specialist Aunty L. Padgham about the project; she gave us permission to use the Taungurung name for the section of the river where this fossil was located (see also Acknowledgements).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Optical surface scans of the footprints shown in Figs. 2c,e and 3a–d can be downloaded via figshare at https://doi.org/10.6084/m9.figshare.25869367 (ref. 57).
References
Marchetti, L. et al. Footprints of the earliest reptiles: Notalacerta missouriensis – ichnotaxonomy, potential trackmakers, biostratigraphy, palaeobiogeography and palaeoecology. Ann. Soc. Geol. Pol. 90, 271–290 (2020).
Marchetti, L. et al. Tracking the origin and early evolution of reptiles. Front. Ecol. Evol. 9, 696511 (2021).
Carroll, R. L. The earliest reptiles. Zool. J. Linn. Soc. 45, 61–83 (1964).
Ford, D. P. & Benson, R. B. J. The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae. Nat. Ecol. Evol. 4, 57–65 (2020).
Milner, A. R. & Sequeira, S. E. K. The temnospondyl amphibians from the Viséan of East Kirkton, West Lothian, Scotland. Earth Environ. Sci. Trans. R. Soc. Edinb. 84, 331–361 (1993).
Pardo, J. D., Szostakiwskyj, M., Ahlberg, P. E. & Anderson, J. S. Hidden morphological diversity among early tetrapods. Nature 546, 642–645 (2017).
Lucas, S. G. et al. in The Carboniferous Timescale Special Publications 512 (eds Lucas, S. G. et al.) 965–1001 (Geological Society, 2021).
Young, G. C. Devonian formations, vertebrate faunas, and age control on the far south coast of New South Wales and adjacent Victoria. Aust. J. Earth Sci. 54, 991–1008 (2007).
Marsden, M. A. H. in Geology of Victoria (eds Douglas, J. G. & Ferguson, J. A.) 147–194 (Victorian Division of the Geological Society of Australia Inc., 1988).
MacDougall, M. J. et al. A new recumbirostran ‘microsaur’ from the Lower Permian Bromacker locality, Thuringia, Germany, and its fossorial adaptations. Sci. Rep. 14, 4200 (2024).
Coates, M. I. & Clack, J. E. Romer’s gap: tetrapod origins and terrestriality. Bull. Mus. Natl Hist. Nat. 17, 373–388 (1995).
Clark, I. D. The Port Phillip Journals of George Augustus Robinson: 8 March-7 April 1842 and 18 March-29 April 1843 (Monash University, Department of Geography, 1988).
Garvey, J. M. & Hasiotis, S. T. An ichnofossil assemblage from the Lower Carboniferous Snowy Plains Formation, Mansfield Basin, Australia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 258, 257–276 (2008).
Garvey, J. M. & Turner, S. Vertebrate microremains from the presumed earliest Carboniferous of the Mansfield Basin, Victoria. Alcheringa 30, 43–62 (2006).
Long, J. A. in Devonian and Carboniferous Fish Studies Memoir 7 (ed. Jell, P. A.) 1–64 (Association of Australasian Palaeontologists, 1988).
Long, J. A. A new rhizodontiform fish from the Early Carboniferous of Victoria, Australia, with remarks on the phylogenetic position of the group. J. Vertebr. Paleontol. 9, 1–17 (1989).
Long, J. A. & Campbell, K. S. W. A new lungfish from the Lower Carboniferous of Victoria, Australia. Proc. R. Soc. Vic. 97, 87–93 (1985).
Fergusson, C. L. & Colquhoun, G. P. Devonian–Carboniferous regional deformation in the northeastern Lachlan Orogen, southeastern Australia. Aust. J. Earth Sci. 68, 1092–1110 (2021).
Foster, D. A. & Gray, D. R. Evolution and structure of the Lachlan Fold Belt (Orogen) of Eastern Australia. Annu. Rev. Earth Planet. Sci. 28, 47–80 (2000).
Berman, D. S., Henrici, A. C., Sumida, S. S. & Martens, T. Redescription of Seymouria sanjuanensis (Seymouriamorpha) from the Lower Permian of Germany based on complete, mature specimens with a discussion of paleoecology of the Bromacker locality assemblage. J. Vertebr. Paleontol. 20, 253–268 (2000).
Kennedy, N. K. Redescription of the postcranial skeleton of Limnoscelis paludis Williston (Diadectomorpha: Limnoscelidae) from the Upper Pennsylvanian of El Cobre Canyon, northern New Mexico. Bull. N. M. Mus. Nat. Hist. Sci. 49, 211–220 (2010).
Marchetti, L., Mujal, E. & Bernardi, M. An unusual Amphisauropus trackway and its implication for understanding seymouriamorph locomotion. Lethaia 50, 162–174 (2016).
Voigt, S., Berman, D. S. & Henrici, A. C. First well-established track-trackmaker association of paleozoic tetrapods based on Ichniotherium trackways and diadectid skeletons from the Lower Permian of Germany. J. Vertebr. Paleontol. 27, 553–570 (2007).
Voigt, S. & Ganzelewski, M. Toward the origin of amniotes: diadectomorph and synapsid footprints from the early Late Carboniferous of Germany. Acta Palaeontol. Pol. 55, 57–72 (2010).
Buchwitz, M., Jansen, M., Renaudie, J., Marchetti, L. & Voigt, S. Evolutionary change in locomotion close to the origin of amniotes inferred from trackway data in an ancestral state reconstruction approach. Front. Ecol. Evol. 9, 674779 (2021).
Maddin, H. C., Eckhart, L., Jaeger, K., Russell, A. P. & Ghannadan, M. The anatomy and development of the claws of Xenopus laevis (Lissamphibi: Anura) reveal alternate pathways of structural evolution in the integument of tetrapods. J. Anat. 214, 607–619 (2009).
Sigurdsen, T. & Bolt, J. R. The Lower Permian amphibamid Doleserpeton (Temnospondyli: Dissorophoidea), the interrelationships of amphibamids, and the origin of modern amphibians. J. Vertebr. Paleontol. 30, 1360–1377 (2010).
Górecka-Nowak, A., Jankowska, A. & Muszer, J. Age revision of Carboniferous rocks in the northern part of the Intra-Sudetic Basin (SW Poland) based on miospore data. Geol. Q. 65, 8 (2021).
Voigt, S., Niedźwiedzki, G., Raczyński, P., Mastalerz, K. & Ptaszyński, T. Early Permian tetrapod ichnofauna from the Intra-Sudetic Basin, SW Poland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 313–314, 173–180 (2012).
Zhu, M., Yu, X. & Ahlberg, P. E. A primitive sarcopterygian fish with an eyestalk. Nature 410, 81–84 (2001).
Lu, J. et al. The earliest known stem tetrapod from the Lower Devonian of China. Nat. Commun. 3, 1160 (2012).
Lu, J. et al. A Devonian predatory fish provides insights into the early evolution of modern sarcopterygians. Sci. Adv. 2, e1600154 (2016).
Niedźwiedzki, G., Szrek, P., Narkiewicz, K., Narkiewicz, M. & Ahlberg, P. E. Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43–48 (2010).
Stössel, I., Williams, E. A. & Higgs, K. T. Ichnology and depositional environment of the Middle Devonian Valentia Island tetrapod trackways, south-west Ireland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 462, 16–40 (2016).
Ahlberg, P. E. Elginerpeton pancheni and the earliest tetrapod clade. Nature 373, 420–425 (1995).
Ahlberg, P. E. Postcranial stem tetrapod remains from the Devonian of Scat Craig, Morayshire, Scotland. Zool. J. Linn. Soc. 122, 99–141 (1998).
Coates, M. I., Ruta, M. & Friedman, M. Ever since Owen: changing perspectives on the early evolution of tetrapods. Annu. Rev. Ecol. Evol. Syst. 39, 571–592 (2008).
Chang, M.-M. & Yu, X.-B. Structure and phylogenetic significance of Diabolepis speratus gen. et sp. nov., a new dipnoan-like form from the Lower Devonian of eastern Yunnan, China. Proc. Linn. Soc. New South Wales 107, 171–184 (1984).
Cloutier, R. et al. Elpistostege and the origin of the vertebrate hand. Nature 579, 549–554 (2020).
Daeschler, E. B., Shubin, N. H. & Jenkins, F. A. Jr. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757–763 (2006).
Shubin, N. H. Your Inner Fish (Vintage, 2009).
Stewart, T. A. et al. The axial skeleton of Tiktaalik rosaeae. Proc. Natl Acad. Sci. USA 121, e2316106121 (2024).
Clack, J. A. Gaining Ground: The Origin and Evolution of Tetrapods 2nd edn (Indiana Univ. Press, 2012).
Ahlberg, P. E. Follow the footprints and mind the gaps: a new look at the origin of tetrapods. Earth Environ. Sci. Trans. R. Soc. Edinb. 109, 115–137 (2019).
Gess, R. & Ahlberg, P. E. A tetrapod fauna from within the Devonian Antarctic Circle. Science 360, 1120–1124 (2018).
Beznosov, P. A., Clack, J. A., Lukševičs, E., Ruta, M. & Ahlberg, P. E. Morphology of the earliest reconstructable tetrapod Parmastega aelidae. Nature 574, 527–531 (2019).
Ahlberg, P. E. & Clack, J. A. The smallest known Devonian tetrapod shows unexpectedly derived features. R. Soc. Open Sci. 7, 192117 (2020).
Camens, A. B. & Carey, S. P. Contemporaneous trace and body fossils from a late Pleistocene lakebed in Victoria, Australia, allow assessment of bias in the fossil record. PLoS ONE 8, e52957 (2013).
Dupret, V. et al. The Bothriolepis (Placodermi, Antiarcha) material from the Valentia Slate Formation of the Iveragh Peninsula (middle Givetian, Ireland): morphology, evolutionary and systematic considerations, phylogenetic and palaeogeographic implications. PLoS ONE 18, e0280208 (2023).
Dupret, V. et al. Non-tetrapod sarcopterygians from the Valentia Slate Formation (Givetian, Devonian) of the Iveragh Peninsula, south-western Ireland: systematic reappraisal and palaeobiogeographic implications. Span. J. Palaeontol. 38, 37–46 (2023).
Klootwijk, C. Sedimentary basins of eastern Australia: paleomagnetic constraints on geodynamic evolution in a global context. Aust. J. Earth Sci. 56, 273–308 (2009).
Klootwijk, C. Australia’s controversial Middle-Late Palaeozoic pole path and Gondwana–Laurasia interaction. Palaeoworld 19, 174–185 (2010).
Clack, J. A. & Finney, S. M. Pederpes finneyae, an articulated tetrapod from the Tournaisian of Western Scotland. J. Syst. Paleontol. 2, 311–346 (2005).
Anderson, J. A., Smithson, T., Mansky, C. F., Meyer, T. & Clack, J. A diverse tetrapod fauna at the base of ‘Romer’s Gap’. PLoS ONE 10, e0125446 (2015).
Clack, J. A. et al. Phylogenetic and environmental context of a Tournaisian tetrapod fauna. Nat. Ecol. Evol. 1, 0002 (2016).
Lebedev, O. A. & Coates, M. I. The postcranial skeleton of the Devonian tetrapod Tulerpeton curtum Lebedev. Zool. J. Linn. Soc. 114, 307–348 (1995).
Long, A. J. et al. Reptile tracks from the earliest Carboniferous of Australia recalibrate the timeline of tetrapod evolution. figshare https://doi.org/10.6084/m9.figshare.25869367 (2025).
Acknowledgements
We acknowledge that NMV P258240 comes from Taungurung Country, and pay our respects to Taungurung Elders past and present, and all of the Taungurung community. We thank Aunty L. Padgham, Taungurung Elder and language specialist, for providing the Taungurung name for the section of the river where this fossil was located; and A. Gołasa, A. Miziołek and P. Menducki for full access to material collected by them from the Sudetic area in Poland, and also for permission to document of the most important specimens from these collections. P.E.A. acknowledges the support of ERC Advanced Grant ERC-2020-ADG 10101963 “Tetrapod Origin”. J.A.L. and A.M.C. receive funding from the Australian Research Council, grants DP 220100825 and DP 200103398.
Funding
Open access funding provided by Uppsala University.
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Jason Anderson, Stuart Sumida and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article
Cite this article
Long, J.A., Niedźwiedzki, G., Garvey, J. et al. Earliest amniote tracks recalibrate the timeline of tetrapod evolution. Nature (2025). https://doi.org/10.1038/s41586-025-08884-5
Received: 21 May 2024
Accepted: 11 March 2025
Published: 14 May 2025
DOI: https://doi.org/10.1038/s41586-025-08884-5
![Mr. Bates vs. the Post Office [TV Series] (Horizon IT Scandal)](https://news.najib.digital/site/assets/img/broken.gif)